Kenaikan CO2 Dan Perubahan Iklim : Implikasinya Terhadap Pertumbuhan Tanaman
Tania June
Ringkasan
Tanaman membutuhkan CO2 untuk pertumbuhannya. Peningkatan konsentrasi CO2 di atmosfir akan merangsang proses fotosintesis, meningkatkan pertumbuhan tanaman dan produktivitas pertanian tanpa diikuti oleh peningkatan kebutuhan air (transpirasi).
Sebaliknya, kenaikan suhu di permukaan bumi mempunyai pengaruh yang "kurang menguntungkan" terhadap pertanian, dapat mengurangi bahkan menghilangkan pengaruh positif dari kenaikan CO2. Tulisan ini merupakan ulasan singkat untuk mengupas hubungan antara kenaikan CO2 dan suhu dengan reaksi yang diberikan tanaman. Pembahasan dititikberatkan pada tingkat daun dengan beberapa contoh-contoh hasil penelitian penulis. Dibahas juga mengenai perbedaan reaksi antara tanaman C3 dan C4 ; perlunya "modelling" untuk memperkirakan hasil pada skala besar dan kompleksnya sistem yang dihadapi.
Perubahan Iklim
Beberapa jenis gas di atmosfir, seperti CO2, CH4, dan N2O mempengaruhi iklim permukaan bumi karena kemampuanya dalam membantu proses transmisi radiasi dari matahari ke permukaan bumi, dan juga menghambat keluarnya sebagian radiasi dari permukaan bumi. Kalau konsentrasi dari gas-gas ini di atmosfir meningkat, radiasi yang keluar dari permukaan bumi akan terhambat, sehingga suhu permukaan bumi bertambah besar. Perkiraan besarnya peningkatan suhu bukanlah pekerjaan yang mudah, karena adanya umpan balik positif (dengan peningkatan uap air , H2O(gas), yang juga merupakan gas penghambat keluarnya radiasi dari permukaan bumi) dan umpan balik negatif (peningkatan pertumbuhan awan, menghambat transmisi radiasi matahari ke permukaan bumi).
Estimasi kenaikan suhu terbaik saat ini adalah yang dihasilkan oleh General Circulation Models (GCM), kenaikan suhu 2.5 - 5.5 oC diikuti dengan kenaikan laju sirkulasi hidrologi sebesar 5-15 % (IPCC, l996).
Pengaruh CO2 terhadap proses fisiologis tanaman
Gas CO2 merupakan sumber karbon utama bagi pertumbuhan tanaman. Konsentrasi CO2 di atmosfir saat ini belum optimal, sehingga penambahan CO2 kepada tanaman di dalam industri pertanian di dalam rumah kaca merupakan kegiatan normal untuk meningkatkan pertumbuhan tanaman seperti tomat, selada, timun dan bunga potong.
Pengaruh fisiologis utama dari kenaikan CO2 adalah meningkatknya laju assimilasi (laju pengikatan CO2 untuk membentuk karbohidrat,fotosintesis) di dalam daun. Efisiensi penggunaan faktor-faktor pertumbuhan lainnya (seperti radiasi matahari, air dan nutrisi) juga akan ikut meningkat.
Proses di tingkat daun
Fotosintesis
Hubungan antara CO2 "ambient" (dapat diartikan sebagai kondisi normal CO2 di atmosfir) dengan proses fotosintesis, baik di tingkat daun maupun di tingkat kanopi tanaman, dan kontribusinya terhadap akumulasi biomasa telah banyak diteliti. Salah satu karya ilmiah yang dengan sangat menarik membahas proses ini adalah "Modelling canopy photosynthesis from the biochemistry of the C3 chloroplast" (Evans and Farquhar, l991).
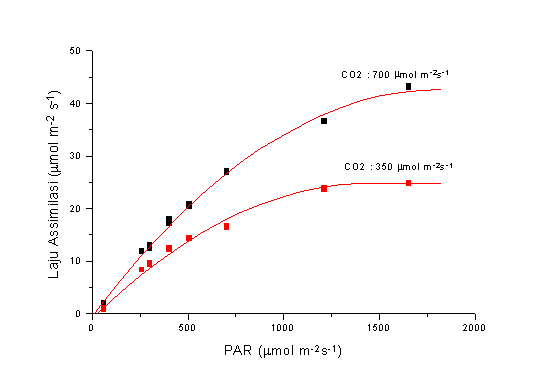
Energi untuk terlaksananya proses fotosintesis datang dari radiasi matahari pada panjang gelombang tertentu (PAR, Photosynthetically Active Radiation, 400-700 m m). Baik PAR, maupun CO2, konsentrasinya masih sub-optimal, sehingga fotosintesis akan meningkat dengan meningkatnya CO2, pada kondisi PAR rendah maupun tinggi.
Untuk lebih jelas, lihat Gambar 1.
Fig. 1. Peningkatan laju assimilasi tanaman kedele (C3) dengan pertambahan PAR
pada konsentarsi CO2 berbeda.
Tanaman terbagi atas dua grup utama, C3 dan C4, yang dibedakan oleh cara mereka mengikat CO2 dari atmosfir dan produk awal yang dihasilkan dari proses assimilasi. Pada tanaman C3, enzim yang menyatukan CO2 dengan RuBP (RuBP merupakan substrat untuk pembentukan karbohidrat dalam proses fotosintesis) dalam proses awal assimilasi, juga dapat mengikat O2 pada saat yang bersamaan untuk proses fotorespirasi ( fotorespirasi adalah respirasi,proses pembongkaran karbohidrat untuk menghasilkan energi dan hasil samping, yang terjadi pada siang hari), sehingga ada kompetisi antara CO2 dan O2 dalam menggunakan RuBP (Farquhar dan Caemmerer, l982). Jika konsentrasi CO2 di atmosfir ditingkatkan, hasil dari kompetisi antara CO2 dan O2 akan lebih menguntungkan CO2, sehingga fotorespirasi terhambat dan assimilasi akan bertambah besar. Contoh tanaman C3 antara lain : kedele, kacang tanah, kentang, sedang contoh tanaman C4 adalah jagung, sorgum dan tebu.
Kalau CO2 di permukaan daun di kurangi, mencapai suatu titik dimana CO2 yang diserap tanaman sama dengan yang dihasilkannya, maka titik ini disebut "CO2 compensation point" (titik kompensasi CO2) (Gambar 2), G . Nilai G adalah sekitar 50 m mol m-2 s-1 pada suhu 25 oC, dan meningkat dengan meningkatnya suhu permukaan daun. Untuk tanaman kedele nilai G lebih rendah, yaitu sekitar 40 m mol m-2 s-1 pada 25 oC.
Nilai ini penting di dalam konteks perubahan iklim yang berkaitan dengan kenaikan suhu. Dengan kenaikan suhu, produksi biomasa akan berkurang jika CO2 di permukaan daun mencapai titik kompensasinya (biasa terjadi di siang hari pada saat matahari terik dan kecepatan angin sangat rendah atau di bawah kanopi hutan tropis), karena G
meningkat. 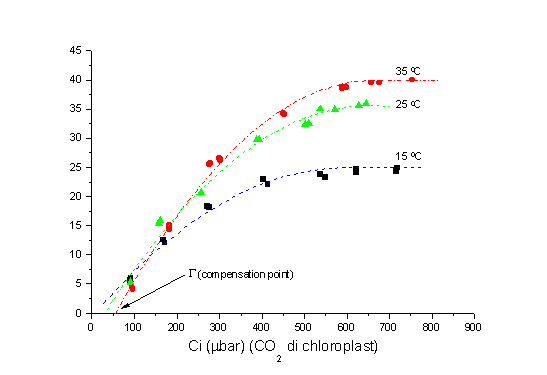
Fig. 2. Laju assimilasi (m mol CO2 m-2 s-1) tanaman kedele dengan meningkatnya
CO2 pada suhu berbeda.
Pada tanaman C4, CO2 diikat oleh PEP (enzym pengikat CO2 pada tanaman C4) yang tidak dapat mengikat O2 sehingga tidak terjadi kompetisi antara CO2 dan O2. Lokasi terjadinya assosiasi awal ini adalah di sel-sel mesofil (sekelompok sel-sel yang mempunyai klorofil yang terletak di bawah sel-sel epidermis daun). CO2 yang sudah terikat oleh PEP kemudian ditransfer ke sel-sel "bundle sheath" (sekelompok sel-sel di sekitar xylem dan phloem) dimana kemudian pengikatan dengan RuBP terjadi. Karena tingginya konsentasi CO2 pada sel-sel bundle sheath ini, maka O2 tidak mendapat kesempatan untuk bereaksi dengan RuBP, sehingga fotorespirasi sangat kecil and G sangat rendah, sekitar 5 m mol m-2 s-1. PEP mempunyai daya ikat yang tinggi terhadap CO2, sehingga reaksi fotosintesis terhadap CO2 di bawah 100 m mol m-2 s-1 sangat tinggi. Pada kisaran konsentrasi CO2 300 - 500 m mol m-2 s-1, laju assimilasi tanaman C4 hanya bertambah sedikit dengan meningkatnya CO2, walaupun
PAR sangat tinggi. Sehingga, dengan meningkatnya CO2 di atmosfir, tanaman C3 akan lebih beruntung dari tanaman C4 dalam hal pemanfaatan CO2 yang berlebihan.
Jika kita kembali ke Gambar 2, terlihat bahwa meningkatnya suhu daun dari 15 oC ke 35 oC menyebabkan laju asimilasi bertambah besar. Meningkatnya asimilasi dengan kenaikan suhu merupakan fenomena umum, sampai suhu optimum tercapai, lalu akan terjadi penurunan, seperti terlihat pada Gambar 3 di bawah ini. Adanya kenaikan CO2 di atmosfir akan merubah suhu optimum tanaman. Untuk tanaman kedele yang saya gunakan, kenaikan suhu optimum mencapai 12 %.
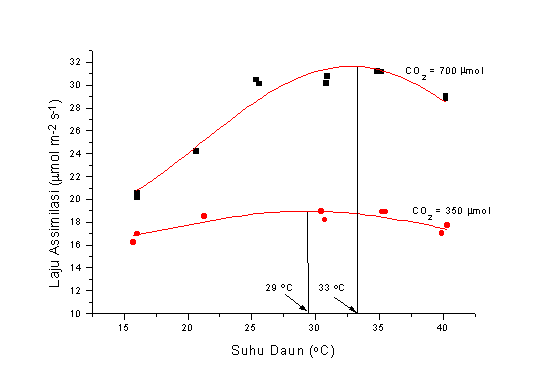
Figure 3. Suhu optimum untuk proses assimilasi akan berubah dengan kenaikan CO2
di atmosfir. Data diambil dari tanaman kedele dan "fitting" menggunakan
persamaan kurva Gauss untuk mendapatkan suhu optimum.
Bertambah besarnya suhu optimum ini menguntungkan bagi tanaman karena pada saat konsentrasi CO2 di atmosfir mencapai 2 kali konsentrasinya saat ini, akan terjadi kenaikan suhu sampai 5.5 oC.
Konduktivitas Stomata
Selain pengaruh positif terhadap proses fotosintesis, kenaikan CO2 juga akan mempunyai pengaruh positif terhadap penggunaan air oleh tanaman.
Stomata mempunyai fungsi sebagai "pintu gerbang" masuknya CO2 dan keluarnya uap air ke/dari daun. Besar kecilnya pembukaan stomata merupakan regulasi terpenting yang dilakukan oleh tanaman, dimana tanaman berusaha memasukkan CO2 sebanyak mungkin tetapi dengan mengeluarkan H2O sesedikit mungkin, untuk mencapai effisiensi pertumbuhan yang tinggi.
Daya ikat yang tinggi terhadap CO2 pada tanaman C4, menyebabkan perbandingan antara pemasukan CO2 dan konduktivitas stomata (kemampuan stomata menyalurkan H2O persatuan waktu) optimum. Dengan kata lain, tanaman-tanaman C4 mempunyai efisiensi penggunaan air yang tinggi. Dengan kata lain, jumlah air yang dikeluarkan untuk sejumlah CO2 yang dimasukkan jauh lebih sedikit pada tanaman C4 dibandingkan dengan tanaman C3. Pada tanaman C3, daya ikat yang rendah terhadap CO2 menyebabkan tanaman ini boros dalam penggunaan air.
Jika CO2 di atmosfir meningkat, tanaman tidak membutuhkan pembukaan stomata maksimum untuk mencapai konsentrasi CO2 optimum di dalam daun, sehingga laju pengeluaran H2O dapat dikurangi. Gambar 3 berikut menunjukkkan bagaimana stomata mengecil dengan meningkatnya CO2.
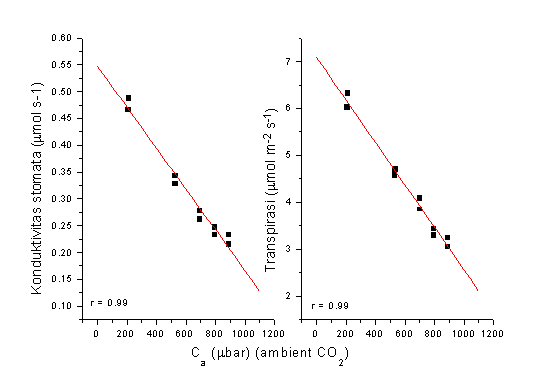
Figure 4. Kenaikan CO2 di atmosfir menyebabkan pembukaan stomata mengecil
(ditunjukkan dengan menurunnya konduktivitas stomata) sehingga
transpirasi dari permukaan daun menurun.
Dari Gambar 4 dapat terlihat bahwa dengan kenaikan CO2 dari 350 ( kondisi normal di atmosfir saat ini) ke700 m bar, konduktivitas stomata menurun sebesar 32 %, menghasilkan penghematan air sebesar 28 %.
Besarnya penurunan ini tergantung dari respon asimilasi tanaman terhadap peningkatan CO2 yang berkisar dari 0 % pada tanaman C4 sampai 40 % pada tanaman C3 (Morison, l987). Efisiensi penggunaan air (ratio antara CO2 yang diikat tanaman dengan jumlah air yang dikeluarkan), baik pada tanaman C3 maupun C4 akan meningkat dengan bertambah besarnya CO2. Pada tanaman C3, peningkatan ini disebabkan oleh meningkatnya asimilasi dan menurunnya transpirasi, sedang pada pada tanaman C4 hanya disebabkan oleh menurunnya transpirasi.
Response pada tingkat Pertanaman
Pertanaman disini saya ambil dari "crop", yaitu sekumpulan tanaman sejenis yang tumbuh berdampingan. Respon pada tingkat pertanaman akan berbeda dari respon di tingkat daun , karena adanya faktor iklim mikro (iklim di sekitar tanaman) yang menyebabkan timbulnya gradien faktor-faktor pertumbuhan dengan kedalaman kanopi, seperti gradien PAR, nitrogen, kecepatan angin, uap air dan CO2. Adanya gradien ini menyebabkan besarnya respon pada tingkat daun akan berkurang pada tingkat pertanaman, apalagi kalau interaksi antara kanopi dengan atmosfir sangat jelek, yang biasa terjadi bila kecepatan angin rendah dan kanopi tanaman menutup permukaan tanah dengan sempurna, sehingga mengurangi distribusi faktor-faktor pertumbuhan ke dalam kanopi.
Rosenberg et al. (l990), dengan menggunakan simulasi iklim mikro, mendapatkan bahwa, penurunan konduktivitas stomata sebesar 40 % di tingkat daun hanya menurunkan penggunaan air sebesar 9 % di tingkat pertanaman. Hal ini mungkin terjadi karena penutupan kanopi yang sempurna menyebabkan gradien kelembaban relatif antara daun dan udara meningkat, sehingga transpirasi bertambah besar.
Proses asimilasi, sebaliknya, mempunyai reaksi yang tidak jauh berbeda dari reaksi di tingkat daun, karena meningkatnya CO2 di atmosfir akan menghilangkan gradien CO2 di kanopi bagian bawah dan meningkatkan asimilasi. Hasilnya, walaupun transpirasi berkurang dibandingkan dengan pengurangan di tingkat daun, asimilasi akan bertambah besar/atau sama dari penambahan di tingkat daun. Kimball (l983) memperoleh 40 % kenaikan biomasa pada tanaman C3 dan 15 % pada tanaman C4.
Modelling
Memperkirakan reaksi pertanaman terhadap kenaikan CO2 merupakan pekerjaan yang tidak mudah mengingat rumitnya sistem yang dihadapi dan terbatasnya pengetahuan kita di dalam mendeteksi interaksi antara mekanisme di dalam tanaman dengan mekanisme di luar tanaman (iklim lapisan perbatas, "boundary layer climate") saat ini. Modelling merupakan salah satu alat mempermudah pekerjaan ini. Keluarnya model mekanistik fotosintesis tanaman C3 di tingkat daun (Farquhar et al., l980; Farquhar and Caemmerer, l982) merupakan suatu terobosan penting di dalam bidang fisiologi tanaman, karena untuk pertama kalinya mekanisme reaksi tanaman (di tingkat daun) terhadap kenaikan CO2, suhu dan PAR secara bersamaan dapat dimengerti. Saya katakan sebagai terobosan penting, karena selama ini perkiraan reaksi tanaman terhadap kenaikan CO2 dan suhu menggunakan model fotosintesis empiris yang kemampuan prediksinya terbatas pada tanaman tertentu dan lokasi tertentu. Dengan keluarnya model mekanistik fotosintesis ini, tingkat kemampuan prediksi akan semakin tinggi. Yang menjadi masalah kemudian adalah "scaling-up" dari tingkat daun ke tingkat pertanaman dan produksi biomasa.
Lloyd et al.(l995) keluar dengan "big-leaf" modelnya, dimana kanopi pertanaman dianggap sebagai sebuah daun besar, sehingga parameter asimilasi dan konduktivitas stomata di tingkat daun dapat digunakan ditingkat pertanaman, sepanjang distribusi PAR di dalam kanopi ikut diperhitungkan. Model ini kemudian digunakan untuk memperkirakan NPP (Net Primary Production, Produksi Bahan Kering Netto) per tahun di hutan hujan tropis di Amazon, dengan hasil yang memuaskan (Lloyd et al., l995).
De Pury and Farquhar (l997) kemudian keluar dengan "Sun/Shade" model, yang memilah kanopi atas dua bagian besar, yaitu bagian yang terkena sinar matahari dan bagian yang terlindungi. Parameter fotosintesis dan konduktifitas stomata dianggap berbeda pada kedua bagian kanopi ini. Sun/Shade model merupakan model yang sangat kompleks, membutuhkan parameter yang lebih banyak dari pada big leaf model. Model ini dapat memprediksi laju fotosintesis dan transpirasi dalam skala waktu yang kecil (detik) di tingkat pertanaman dengan sangat akurat, dibandingkan dengan big leaf model. Akan tetapi prediksi biomasa (dengan skala waktu yang lebih besar, mingguan atau tahunan) dengan menggunakan model ini belum pernah dilakukan.
Menurut Goudriaan (l997, personal communication), guru dalam bidang modelling pertumbuhan tanaman (dari Wageningen University), kompleks model yang memilah-milah kanopi atas beberapa bagian merupakan alat yang penting untuk mempelajari mekanisme fotosintesis kanopi dalam skala waktu kecil, tetapi tidak praktis untuk digunakan meng-estimasi pertumbuhan atau produksi karena besarnya input parameter yang dibutuhkan. Goudriaan (l997, unpublished data), telah mencoba membandingkan estimasi produksi dari big leaf dan "multiple layer " model (model yang membagi kanopi atas beberapa bagian vertikal, kemudian memilah setiap bagian atas dua bagian yaitu bagian tersinari dan terlindungi ) dan mendapatkan hasil yang tidak berbeda, walaupun variasi fotosintesis diurnal menunjukkan bahwa multiple layer model lebih akurat. Hal ini mungkin terjadi karena perbedaan estimasi fotosintesis di tingkat kanopi tidak cukup besar untuk menimbulkan perbedaan estimasi produksi. Selain itu, produksi akhir tanaman tidak hanya ditentukan oleh besarnya fotosintesis/asimilasi ; partisi hasil fotosintesis ke bagian-bagian tanaman merupakan faktor sangat penting yang penguasaan mekanisme prosesnya tidak sebaik penguasaan kita terhadap proses fotosintesis.
Pemilihan model yang akan digunakan dalam prediksi respon tanaman terhadap CO2, tergantung dari seberapa jauh detail dari jawaban yang kita inginkan. Kalau hanya tertarik pada produksi akhir (dan mekanisme diurnal yang cukup akurat), maka big leaf model merupakan pilihan yang tepat, karena lebih sederhana, membutuhkan parameter lebih sedikit sehingga pembuatan "source code" untuk simulasi tidak terlalu sulit. Akan tetapi kalau kita lebih tertarik dengan mekanisme diurnal yang sangat akurat, Sun/Shade model atau Multiple Layer model merupakan pilihan yang tepat, walaupun membutuhkan penelitian awal yang intensif terhadap tanaman yang akan diteliti untuk memperoleh parameter yang dibutuhkan, ditambah lagi rumitnya persamaan matematik yang digunakan membutuhkan kemampuan programming yang cukup baik untuk dapat membuat source code yang jelas.
Kesimpulan
Bahasan di dalam tulisan ini masih sangat kurang untuk dapat mengupas secara lengkap pengaruh perubahan iklim terhadap produktivitas pertanian, baik secara langsung (meningkatnya CO2) maupun secara tidak langsung (meningkatnya suhu, berubahnya keawanan dan curah hujan), karena tulisan ini lebih menitik beratkan kepada pengaruh perubahan iklim terhadap proses fisiologis tanaman dalam skala spatial yang relatif kecil. Untuk melihat pengaruhnya secara regional dan global, maka faktor- faktor penting lainnya, seperti: 1) bagaimana sirkulasi iklim umum global berubah, yang akan menentukan perubahan jumlah hujan yang diterima ditiap region dan besarnya "regional temperature shifting". Regional temperature shifting adalah bergeraknya zona temperature ke arah kutub, yang mengakibatkan semakin luasnya daerah tropis dan mengecilnya daerah kutub. Kalau pergerakan ini terjadi maka daerah yang biasanya bukan daerah pertanian produktif, karena suhu terlalu rendah, akan menjadi daerah pertanian baru. Akan tetapi, untuk daerah-daerah yang keadaan suhunya sudah optimum untuk pertumbuhan tanaman, dengan adanya kenaikan suhu, akan mengurangi fotosintesis dan mempercepat perkembangan tanaman sehingga produksi akan turun; 2) berpindahnya hama dan penyakit tanaman dari satu daerah ke daerah lainnya, atau berubahnya derajat penyerangan dengan berubahnya kondisi iklim;
3) perbedaan kesuburan tanah secara regional, yang akan menentukan bagaimana tanaman bereaksi terhadap kenaikan CO2, suhu dan kemungkinan perubahan curah hujan; 4) kemungkinan mencairnya es di kutub yang yang menaikkan permukaan air laut. IPCC (Intergovernmental Panel on Climate Change) memprediksi adanya kenaikan air laut sebesar 28-96 cm pada tahun 2090. Asia tenggara akan mendapat ancaman banjir terbesar karena umumnya, lahan pertanian di region ini terletak pada altitut (ketinggian) rendah (IPCC, l996); 5) masih kurangnya kemampuan GCM (General Circulation Model) dalam memprediksi secara akurat seberapa besar perubahan iklim per daerah yang akan dan mungkin terjadi dan 6) masih kurangnya pengetahuan kita terhadap kemungkinan adaptasi atau aklimasi yang mungkin dilakukan oleh alam (terutama tanaman) sehingga memperkecil atau bahkan menghilangkan perubahan iklim.
Masih banyak penelitian yang harus dilakukan untuk mempersiapkan pertanian kita terhadap kemungkinan terburuk dari perubahan iklim, baik dengan melakukan penelitian di lapang terhadap berbagai jenis tanaman pertanian penting maupun dengan penyempurnaan model-model pertumbuhan tanaman yang ada, dan penyempurnaan GCM sehingga kita siap menghadapinya kalau dan bila dia datang.
Bibliografi
De Pury, D. G. G. and Farquhar, G. D. l997. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf model. Plant, Cell and Environment. 20: 537-557.
Evans, J. R. and Farquhar, G. D. l991. Modelling canopy photosynthesis from the biochemistry of the C3 Chloroplast. In Boote, K. J. and Loomis, R. S. (eds.) Modelling crop photosynthesis-from biochemistry to canopy. CSSA Special Publication no. l9. American Society of Agronomy and Crop Science Society of America. Madison.
Farquhar, G. D. and von Caemmerer, S. l982. Modelling of photosynthesis response to environmental conditions. p. 550-587. In O.L. Lange et al. (ed.) Encycl. Plant Physiol. New Ser. Vol. 12B. Springer-Verlag, Berlin.
Farquhar, G. D., von Caemmerer, S., and Berry, J. A. l980. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta. 149:78-90.
IPCC. l996. Climate change l995 : The science of climate change. Cambridge University Press.
Lloyd, J., Wong, S. C., Styles, J. M., Batten, D., Priddle, R., Turnbull, C., and McConchie, C. A. l995. Measuring and modelling whole-tree gas exchange. Aust. J. Plant Physiology. 22 : 987-1000.
Lloyd, J., Grace, J., Miranda, A. C., Meir, P., Wong, S. C., Miranda, H. S., Wright, I. R., Gash, J. H. C. and McIntyre, J. l995. A simple calibrated model of Amazon rainforest productivity based on leaf biochemical properties. Plant, Cell and Environment. 18: 1129-1145.
Morison, J. I. L.. l987. Intercellular CO2 concentration and irradiance on the stomatal response to CO2. In E. Zeiger et al.,(ed.) Stomatal function. Stanford Univ. Press. California.
Kimball, B. A.. l983. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agron. J. 75:779-788.
Rosenberg, N. J., Kimball, B. A., Martin, Ph. and Cooper, C. F. l990. From climate and CO2 enrichment to evapotranspiration. p. 151-175. In P. E. Waggoner (ed.) Climate change and US water resources. John Wiley & Sons, New York.